Cellular Organization
Although there are many smaller units such as enzymes and organelles that perform physiological tasks or have definable structures, the smallest anatomical and physiological unit in the human body that can, under appropriate conditions, live and reproduce on its own is the cell. Cells were first discovered more than 300 years ago shortly after Antony van Leeuwenhoek, a Dutch optician, invented the microscope. With his microscope, van Leeuwenhoek was able to observe “many very small animalcules, the motions of which were very pleasing to behold” in tartar scrapings from his teeth. Following the efforts of van Leeuwenhoek, Robert Hooke, a Curator of Instruments for the Royal Society of England, in the late 1600s further described cells when he used one of the earliest microscopes to look at the plant cell walls that remain in cork. These observations and others led to the cell theory developed by Theodor Schwann and Matthias Jakob Schleiden and formalized by Rudolf Virchow in the mid-1800s. The cell theory states that (1) all organisms are composed of one or more cells, (2) the cell is the smallest unit of life, and (3) all cells come from previously existing cells. Thus, cells are the basic building blocks of life.
Cells are composed mostly of organic compounds and water, with more than 60 percent of the weight in a human body coming from water. The organic compounds—carbohydrates, lipids, proteins, and nucleic acids—that cells synthesize are the molecules that are fundamental to sustaining life. These molecules function as energy packets, storehouses of energy and hereditary information, structural materials, and metabolic workers. The most common elements found in humans (in descending order based on percent of body weight) are oxygen, carbon, hydrogen, nitrogen, calcium, phosphorus, potassium, sodium, chlorine, magnesium, sulfur, iron, and iodine. Carbon, hydrogen, oxygen, and nitrogen contribute more than 99 percent of all the atoms in the body. Most of these elements are incorporated into organic compounds, but some exist in other forms, such as phosphate groups and ions.
Carbohydrates are used by cells not only as structural materials but also to transport and store energy. The three classes of carbohydrates are monosaccharides (e.g., glucose), oligosaccharides (e.g., lactose, sucrose, maltose), and polysaccharides (e.g., glycogen). Lipids are greasy or oily compounds that will dissolve in each other but not in water. They form structural materials in cells and are the main reservoirs of stored energy. Proteins are the most diverse form of biological molecules. Specialized proteins, called enzymes, make metabolic reactions proceed at a faster rate than would occur if the enzymes were not available and enable cells to produce the organic compounds of life. Other proteins provide structural elements in the body, act as transport channels across plasma membranes, function as signals for changing activities, and provide chemical weapons against disease-carrying bacteria. These diverse proteins are built from a small number (20) of essential amino acids.
Nucleotides and nucleic acids make up the last category of important biological molecules. Nucleotides are small organic compounds that contain a five-carbon sugar (ribose or deoxyribose), a phosphate group, and a nitrogen-containing base that has a single or double carbon ring structure. Adenosine triphosphate (ATP) is the energy currency of the cell and plays a central role in metabolism. Other nucleotides are subunits of coenzymes that are enzyme helpers. The two nucleic acids are deoxyribonucleic acid (DNA) and ribonucleic acid (RNA). DNA (Figure 3.3) is a unique, helical molecule that contains chains of paired nucleotides that run in opposite directions. Each nucleotide contains either a pyrimidine base—thymine (T) or cytosine (C)—with a single ring structure or a purine base— adenine (A) or guanine (G)—with a double ring. In the double helix of DNA, thymine always pairs with adenine (T-A) and cytosine always pairs with guanine (C-G). RNA is similar to DNA except that it consists of a single helical strand, contains ribose instead of deoxyribose, and has uracil (U) instead of thymine.
All cells are surrounded by a plasma membrane that separates, but does not isolate, the cell’s interior from its environment. Animal cells, such as those found in humans, are eukaryotic cells. A generalized animal cell is shown in Figure 3.4. In addition to the plasma membrane, eukaryotic cells contain membrane-bound organelles and a membrane-bound nucleus. Prokaryotic cells, such as bacteria, lack membrane-bound structures other than the plasma membrane. In addition to a plasma membrane, all cells have a region that contains DNA (which carries the hereditary instructions for the cell) and cytoplasm (which is a semifluid substance that includes everything inside the plasma membrane except for the DNA).
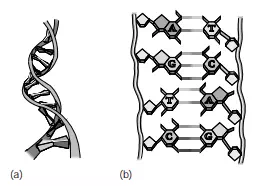
FIGURE 3.3 (a) DNA consists of two chains of paired nucleotides that run in opposite directions and form a helical structure. (b) Thymine pairs with adenine (T-A) and cytosine pairs with guanine (C-G) due to hydrogen bonding between the bases.
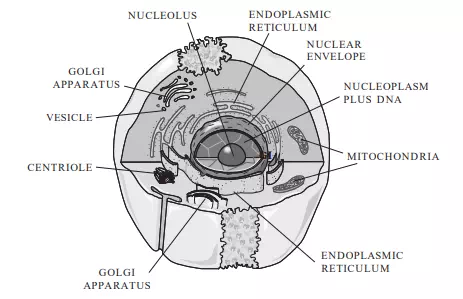
FIGURE 3.4 Animal cells are surrounded by a plasma membrane. They contain a membrane-bound region, the nucleus, which contains DNA. The cytoplasm lies outside of the nucleus and contains several types of organelles that perform specialized functions.
Plasma Membrane
The plasma membrane performs several functions for the cell. It gives mechanical strength, provides structure, helps with movement, and controls the cell’s volume and its activities by regulating the movement of chemicals in and out of the cell. The plasma membrane is composed of two layers of phospholipids interspersed with proteins and cholesterol (Figure 3.5). The proteins in the plasma membranes of mammalian cells provide
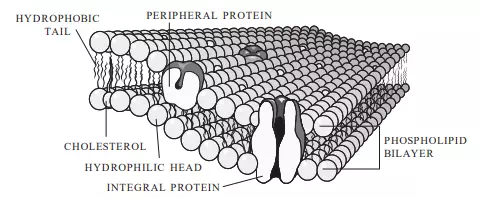
FIGURE 3.5 The plasma membrane surrounds all cells. It consists of a double layer of phospholipids interspersed with proteins and cholesterol.
binding sites for hormones, recognition markers for identifying cells as one type or another, adhesive mechanisms for binding adjacent cells to each other, and channels for transporting materials across the plasma membrane. The phospholipids are arranged with their “water loving” (hydrophilic) heads pointing outward and their “water fearing” (hydrophobic) tails pointing inward. This double-layer arrangement of phospholipids interspersed with protein channels helps maintain the internal environment of a cell by controlling the substances that move across the membrane, whereas the cholesterol molecules act as stabilizers to prevent extensive lateral movement of the lipid molecules. Some molecules, such as oxygen, carbon dioxide, and water, can easily cross the plasma membrane, whereas other substances, such as large molecules and ions, must move through the protein channels. Osmosis is the process by which substances move across a selectively permeable membrane such as a cell’s plasma membrane, whereas diffusion refers to the movement of molecules from an area of relatively high concentration to an area of relatively low concentration. Substances that can easily cross the plasma membrane achieve diffusion equilibrium when there is no net movement of these substances across the membrane; that is, the concentration of the substance inside the cell equals the concentration of the substance outside of the cell. Active transport, which requires an input of energy usually in the form of ATP, can be used to move ions and molecules across the plasma membrane and is often used to move them from areas of low concentration to areas of high concentration. This mechanism helps maintain concentrations of ions and molecules inside a cell that are different from the concentrations outside the cell. A typical mammalian cell has internal sodium ion (Na+) concentrations of 12 mM (12 moles of Na+ per 1,000 litters of solution) and extracellular Na+ concentrations of 120 mM, whereas intracellular and extracellular potassium ion (K+) concentrations are on the order of 125 mM and 5 mM, respectively. In addition to positively charged ions (cations), cells also contain negatively charged ions (anions). A typical mammalian cell has intracellular and extracellular chloride ion (Cl) concentrations of 5 mM and 125 mM and internal anion (e.g., proteins, charged amino acids, sulphate ions, and phosphate ions) concentrations of 108 mm These transmembrane ion gradients are used to make ATP, to drive various transport processes, and to generate electrical signals.
The plasma membrane plays an important role in regulating cell volume by controlling the internal osmolarity of the cell. Osmolarity is defined in terms of concentration of dissolved substances. A 1 osmolar (1 Osm) solution contains 1 mole of dissolved particles per litter of solution, while a 1 milliosmolar (1 mOsm) solution has 1 mole of dissolved particles per 1,000 litters of solution. Thus, solutions with high osmolarity have low concentrations of water or other solvents. For biological purposes, solutions with 0.1 Osm glucose and 0.1 Osm urea have essentially the same concentrations of water. It is important to note that a 0.1 M solution of sodium chloride (NaCl) will form a 0.2 Osm solution, since NaCl dissociates into Na+ and Cl ions and thus has twice as many dissolved particles as a solution of a substance—for example, glucose—that does not dissociate into smaller units. Two solutions are isotonic if they have the same osmolarity. One solution is hypotonic to another if it has a lower osmolarity and hypertonic to another if it has a higher osmolarity. It is important to note that tonicity (isotonic, hypotonic, or hypertonic) is only determined by those molecules that cannot cross the plasma membrane, since molecules that can freely cross will eventually reach equilibrium with the same concentration inside and outside of the cell.
Consider a simple model cell that consists of a plasma membrane and cytoplasm. The cytoplasm in this model cell contains proteins that cannot cross the plasma membrane and water that can. At equilibrium, the total osmolarity inside the cell must equal the total osmolarity outside the cell. If the osmolarity inside and the osmolarity outside of the cell are out of balance, there will be a net movement of water from the side of the plasma membrane where it is more highly concentrated to the other until equilibrium is achieved. For example, assume that a model cell (Figure 3.6) contains 0.2 M protein and is placed in a hypotonic solution that contains 0.1 M sucrose. The plasma membrane of this model cell is impermeable to proteins and sucrose but freely permeable to water. The volume of the

FIGURE 3.6 A simple model cell that consists of cytoplasm, containing 0.2 M proteins, and a plasma membrane is placed in a solution of 0.1 M sucrose. The plasma membrane is insoluble to proteins and sucrose but allows water to pass freely in either direction. The full extent of the extracellular volume is not shown and is much larger than the cell’s volume of 1 nl.
cell, 1 nl, is very small relative to the volume of the solution. In other words, changes in the cell’s volume have no measurable effect on the volume of the external solution. What will happen to the volume of the cell as it achieves equilibrium? At equilibrium, the osmolarity inside the cell must equal the osmolarity outside the cell. The initial osmolarity inside the cell is 0.2 Osm, since the proteins do not dissociate into smaller units. The osmolarity outside the cell is 0.1 Osm due to the sucrose solution. A 0.2 Osm solution has 0.2 moles of dissolved particles per litre of solution, while a 0.1 Osm solution has half as many moles of dissolved particles per litre. The osmolarity inside the cell must decrease by a factor of 2 in order to achieve equilibrium. Since the plasma membrane will not allow any of the protein molecules to leave the cell, this can only be achieved by doubling the cell’s volume. Thus, there will be a net movement of water across the plasma membrane until the cell’s volume increases to 2 nl and the cell’s internal osmolarity is reduced to 0.1 Osm—the same as the osmolarity of the external solution. The water moves down its concentration gradient by diffusing from where it is more highly concentrated in the 0.1 M sucrose solution to where it is less concentrated in the 0.2 M protein solution in the cell.
Real cells are much more complex than the simple model just described. In addition to achieving osmotic balance at equilibrium, real cells must also achieve electrical balance with regard to the ions that are present in the cytoplasm. The principle of electrical neutrality requires that the overall concentration of cations in a biological compartment—for example, a cell—must equal the overall concentration of anions in that compartment. Consider another model cell (Figure 3.7) with internal and external cation and anion concentrations similar to those of a typical mammalian cell. Is the cell at equilibrium if the plasma membrane is freely permeable to K+ and Cl but impermeable to Na+ and the internal anions? The total osmolarity inside the cell is 250 mOsm (12 mM Na+, 125 mM K+, 5 mM Cl, 108 mM anions), while the total osmolarity outside the cell is also 250 mOsm (120 mM Na+, 5 mM K+, 125 mM Cl), so the cell is in osmotic balance—that is, there will be no net movement of water across the plasma membrane. If the average charge per molecule of the anions inside the cell is considered to be –1.2, then the cell is also approximately in electrical equilibrium (12 þ 125 positive charges for Na+ and K+; 5 þ 1.2 * 108 negative charges for Cl and the other anions). Real cells, however, cannot maintain this equilibrium without expending energy, since real cells are slightly permeable to Na+. In order to maintain equilibrium and keep Na+ from accumulating intracellularly, mammalian cells must actively pump Na+ out of the cell against its diffusion and electrical gradients. Since Na+
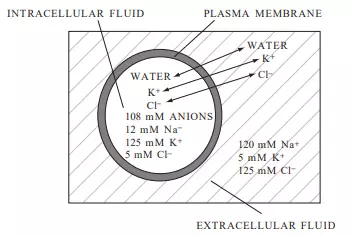
FIGURE 3.7 A model cell with internal and external concentrations similar to those of a typical mammalian cell. The full extent of the extracellular volume is not shown and is much larger than the cell’s volume.
is pumped out through specialized protein channels at a rate equivalent to the rate at which it leaks in through other channels, it behaves osmotically as if it cannot cross the plasma membrane. Thus, mammalian cells exist in a steady state, rather than at equilibrium, since energy in the form of ATP must be used to prevent a net movement of ions across the plasma membrane One of the consequences of the distribution of charged particles in the intracellular and extracellular fluids is that an electrical potential exists across the plasma membrane. The value of this electrical potential depends on the intracellular and extracellular concentrations of ions that can cross the membrane and will be described more fully in Chapter 11.
In addition to controlling the cell’s volume, the plasma membrane also provides a route for moving large molecules and other materials into and out of the cell. Substances can be moved into the cell by means of endocytosis (Figure 3.8a) and out of the cell by means of exocytosis (Figure 3.8b). In endocytosis, material—for example, a bacterium—outside of the cell is engulfed by a portion of the plasma membrane that encircles it to form a vesicle. The vesicle then pinches off from the plasma membrane and moves its contents to the inside of the cell. In exocytosis, material within the cell is surrounded by a membrane to form a vesicle. The vesicle then moves to the edge of the cell, where its membrane fuses with the plasma membrane and its contents are released to the exterior of the cell.

FIGURE 3.8 Substances that are too large to pass through the integral proteins in the plasma membrane can be moved into the cell by means of endocytosis (a) and out of the cell by means of exocytosis (b).