Nutrients of Human Metabolism
Carbohydrates, lipids, and proteins are the major constituents of foods and serve as fuel molecules for the human body. The digestion (breaking down into smaller pieces) of these nutrients in the alimentary tract and the subsequent absorption (entry into the bloodstream) of the digestive end products make it possible for tissues and cells to transform the potential chemical energy of food into useful work.
The major absorbed end products of food digestion are monosaccharides, mainly glucose (from carbohydrates); monoacylglycerol and long-chain fatty acids (from lipids); and small peptides and amino acids (from protein). Once in the bloodstream, different cells can metabolize these nutrients. We have long known that these three classes of molecules are fuel sources for human metabolism, yet it is a common misconception (especially among undergraduates) that human cells use only glucose as a source of energy. This misinformation may arise from the way most textbooks explain energy metabolism, emphasizing glycolysis (the metabolic pathway for glucose degradation) and omitting fatty acid or amino acid oxidation. Here we discuss how the three nutrients (carbohydrates, proteins, and lipids) are metabolized in human cells in a way that may help avoid this oversimplified view of the metabolism.
Historical Overview of Energy Metabolism
During the eighteenth century, the initial studies, developed by Joseph Black, Joseph Priestley, Carl Wilhelm Scheele, and Antoine Lavoisier, played a special role in identifying two gases, oxygen and carbon dioxide, that are central to energy metabolism. Lavoisier, the French nobleman who owns the title of "father of modern chemistry," characterized the composition of the air we breathe and conducted the first experiments on energy conservation and transformation in the organism.
One of Lavoisier's main questions at this time was: How does oxygen's role in combustion relate to the process of respiration in living organisms? Using a calorimeter to make quantitative measurements with guinea pigs and later on with himself and his assistant, he demonstrated that respiration is a slow form of combustion (Figure 1). Based on the concept that oxygen burned the carbon in food, Lavoisier showed that the exhaled air contained carbon dioxide, which was formed from the reaction between oxygen (present in the air) and organic molecules inside the organism. Lavoisier also observed that heat is continually produced by the body during respiration. It was then, in the middle of the nineteenth century, that Justus Liebig conducted animal studies and recognized that proteins, carbohydrates, and fats were oxidized in the body. Finally, pioneering contributions to metabolism and nutrition came from the studies of a Liebig's protégé, Carl von Voit, and his talented student, Max Rubner. Voit demonstrated that oxygen consumption is the result of cellular metabolism, while Rubner measured the major energy value of certain foods in order to calculate the caloric values that are still used today. For example, carbohydrates and proteins produce approximately 4 kcal/g of energy, whereas lipids can generate up to 9 kcal/g. Rubner's observations proved that, for a resting animal, heat production was equivalent to heat elimination, confirming that the law of conservation of energy, implied in Lavoisier's early experiments, was applicable to living organisms as well. Therefore, what makes life possible is the transformation of the potential chemical energy of fuel molecules through a series of reactions within a cell, enabled by oxygen, into other forms of chemical energy, motion energy, kinetic energy, and thermal energy.
Energy Conservation: Mechanisms of ATP Synthesis
Energy metabolism is the general process by which living cells acquire and use the energy needed to stay alive, to grow, and to reproduce. How is the energy released while breaking the chemical bonds of nutrient molecules captured for other uses by the cells? The answer lies in the coupling between the oxidation of nutrients and the synthesis of high-energy compounds, particularly ATP, which works as the main chemical energy carrier in all cells.
There are two mechanisms of ATP synthesis: 1. oxidative phosphorylation, the process by which ATP is synthesized from ADP and inorganic phosphate (Pi) that takes place in mitochondrion; and 2. substrate-level phosphorylation, in which ATP is synthesized through the transfer of high-energy phosphoryl groups from high-energy compounds to ADP. The latter occurs in both the mitochondrion, during the tricarboxylic acid (TCA) cycle, and in the cytoplasm, during glycolysis. In the next section, we focus on oxidative phosphorylation, the main mechanism of ATP synthesis in most of human cells. Later we comment on the metabolic pathways in which the three classes of nutrient molecules are degraded
Oxidative Phosphorylation: The Main Mechanism of ATP Synthesis in Most Human Cells
Figure Detail
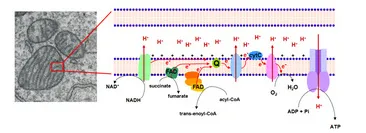
The metabolic reactions are energy-transducing processes in which the oxidation-reduction reactions are vital for ATP synthesis. In these reactions, the electrons removed by the oxidation of fuel molecules are transferred to two major electron carrier coenzymes, nicotinamide adenine dinucleotide (NAD+) and flavin adenine dinucleotide (FAD), that are converted to their reduced forms, NADH and FADH2. Oxidative phosphorylation depends on the electron transport from NADH or FADH2 to O2, forming H2O. The electrons are "transported" through a number of protein complexes located in the inner mitochondrial membrane, which contains attached chemical groups (flavins, iron-sulfur groups, heme, and cooper ions) capable of accepting or donating one or more electrons (Figure 2). These protein complexes, known as the electron transfer system (ETS), allow distribution of the free energy between the reduced coenzymes and the O2 and more efficient energy conservation.
The electrons are transferred from NADH to O2 through three protein complexes: NADH dehydrogenase, cytochrome reductase, and cytochrome oxidase. Electron transport between the complexes occurs through other mobile electron carriers, ubiquinone and cytochrome c. FAD is linked to the enzyme succinate dehydrogenase of the TCA cycle and another enzyme, acyl-CoA dehydrogenase of the fatty acid oxidation pathway. During the reactions catalyzed by these enzymes, FAD is reduced to FADH2, whose electrons are then transferred to O2 through cytochrome reductase and cytochrome oxidase, as described for NADH dehydrogenase electrons (Figure 2).
The electron transfer through the components of ETS is associated with proton (H+) pumping from the mitochondrial matrix to intermembrane space of the mitochondria. These observations led Peter Mitchell, in 1961, to propose his revolutionary chemiosmotic hypothesis. In this hypothesis, Mitchell proposed that H+ pumping generates what he called the proton motive force, a combination of the pH gradient across the inner mitochondrial membrane and the transmembrane electrical potential, which drives the ATP synthesis from ADP and Pi. ATP is synthesized by the ATP synthase complex, through which H+ protons return to the mitchondrial matrix (Figure 2, far right). Paul Boyer first described the ATP synthase catalytic mechanism and showed both that the energy input from the H+ gradient was used for ATP release from the catalytic site, and that the three active sites of the enzyme worked cooperatively in such a way that ATP from one site could not be released unless ADP and Pi were available to bind to another site.
Oxidation of Carbohydrates, Proteins, and Fats Converge on the Tricarboxylic Acid Cycle
Figure 3: Reactions of tricarboxylic acid cycle
The reactions catalyzed by the dehydrogenases that result in NAD+ and FAD reduction are highlighted. The reaction catalyzed by succinyl-CoA synthetase (in which GTP synthesis occurs) is an example of substrate-level phosphorylation.
Interconversion of energy between reduced coenzymes and O2 directs ATP synthesis, but how (and where) are NADH and FADH2 reduced? In aerobic respiration or aerobiosis, all products of nutrients' degradation converge to a central pathway in the metabolism, the TCA cycle. In this pathway, the acetyl group of acetyl-CoA resulting from the catabolism of glucose, fatty acids, and some amino acids is completely oxidized to CO2 with concomitant reduction of electron transporting coenzymes (NADH and FADH2). Consisting of eight reactions, the cycle starts with condensing acetyl-CoA and oxaloacetate to generate citrate (Figure 3). The next seven reactions regenerate oxaloacetate and include four oxidation reactions in which energy is conserved with the reduction of NAD+ and FAD coenzymes to NADH and FADH2, whose electrons will then be transferred to O2 through the ETS. In addition, a GTP or an ATP molecule is directly formed as an example of substrate-level phosphorylation. In this case, the hydrolysis of the thioester bond of succinyl-CoA with concomitant enzyme phosphorylation is coupled to the transfer of an enzyme-bound phosphate group to GDP or ADP. Importantly, although O2 does not participate directly in this pathway, the TCA cycle only operates in aerobic conditions because the oxidized NAD+ and FAD are regenerated only in the ETS. Also noteworthy is that TCA cycle intermediates may also be used as the precursors of different biosynthetic processes.
The TCA cycle is also known as the Krebs cycle, named after its discoverer, Sir Hans Kreb. Krebs based his conception of this cycle on four main observations made in the 1930s. The first was the discovery in 1935 of the sequence of reactions from succinate to fumarate to malate to oxaloacetate by Albert Szent-Gyorgyi, who showed that these dicarboxylic acids present in animal tissues stimulate O2 consumption. The second was the finding of the sequence from citrate to α-ketoglutarate to succinate, in 1937, by Carl Martius and Franz Knoop. Next was the observation by Krebs himself, working on muscle slice cultures, that the addition of tricarboxylic acids even in very low concentrations promoted the oxidation of a much higher amount of pyruvate, suggesting a catalytic effect of these compounds. And the fourth was Krebs's observation that malonate, an inhibitor of succinate dehydrogenase, completely stopped the oxidation of pyruvate by the addition of tricarboxylic acids and that the addition of oxaloacetate in the medium in this condition generated citrate, which accumulated, thus elegantly showing the cyclic nature of the pathway.
Pathways for Nutrient Degradation that Converge onto the TCA Cycle
Glycolysis
Figure 4
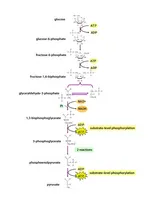
Glycolysis is the pathway in which one glucose molecule is degraded into two pyruvate molecules. Interestingly, during the initial phase, energy is consumed because two ATP molecules are used up to activate glucose and fructose-6-phosphate. Part of the energy derived from the breakdown of the phosphoanhydride bond of ATP is conserved in the formation of phosphate-ester bonds in glucose-6-phosphate and fructose-1,6-biphosphate (Figure 4).
In the second part of glycolysis, the majority of the free energy obtained from the oxidation of the aldehyde group of glyceraldehyde 3-phosphate (G3P) is conserved in the acyl-phosphate group of 1,3- bisphosphoglycerate (1,3-BPG), which contains high free energy. Then, part of the potential energy of 1,3BPG, released during its conversion to 3-phosphoglycerate, is coupled to the phosphorylation of ADP to ATP. The second reaction where ATP synthesis occurs is the conversion of phosphoenolpyruvate (PEP) to pyruvate. PEP is a high-energy compound due to its phosphate-ester bond, and therefore the conversion reaction of PEP to pyruvate is coupled with ADP phosphorylation. This mechanism of ATP synthesis is called substrate-level phosphorylation.
For complete oxidation, pyruvate molecules generated in glycolysis are transported to the mitochondrial matrix to be converted into acetyl-CoA in a reaction catalyzed by the multienzyme complex pyruvate dehydrogenase (Figure 5). When Krebs proposed the TCA cycle in 1937, he thought that citrate was synthesized from oxaloacetate and pyruvate (or a derivative of it). Only after Lipmann's discovery of coenzyme A in 1945 and the subsequent work of R. Stern, S. Ochoa, and F. Lynen did it become clear that the molecule acetyl-CoA donated its acetyl group to oxaloacetate. Until this time, the TCA cycle was seen as a pathway to carbohydrate oxidation only. Most high school textbooks reflect this period of biochemistry knowledge and do not emphasize how the lipid and amino acid degradation pathways converge on the TCA cycle.
The Fatty Acid Oxidation Pathway Intersects the TCA Cycle
Figure 5
In 1904, Knoop, in a classic experiment, decisively showed that fatty acid oxidation was a process by which two-carbon units were progressively removed from the carboxyl end fatty acid molecule. The process consists of four reactions and generates acetyl-CoA and the acyl-CoA molecule shortened by two carbons, with the concomitant reduction of FAD by enzyme acyl-CoA dehydrogenase and of NAD+ by β-hydroxyacyl-CoA dehydrogenase. This pathway is known as β-oxidation because the β-carbon atom is oxidized prior to when the bond between carbons β and α is cleaved (Figure 6). The four steps of β-oxidation are continuously repeated until the acyl-CoA is entirely oxidized to acetyl-CoA, which then enters the TCA cycle. In the 1950s, a series of experiments verified that the carbon atoms of fatty acids were the same ones that appeared in the acids of TCA cycle.
Amino Acid Transamination/Deamination Contributes to the TCA Cycle
Two points must be considered regarding the use of amino acids as fuels in energy metabolism. The first is the presence of nitrogen in amino acid composition, which must be removed before amino acids become metabolically useful. The other is that there are at least twenty different amino acids, each of which requires a different degradation pathway. For our purpose here, it is important to mention two kinds of reactions involving amino acid: transamination and deamination. In the first kind of reaction, the enzymes aminotransferases convert amino acids to their respective α-ketoacids by transferring the amino group of one amino acid to an α-ketoacid. This reaction allows the amino acids to be interconverted. The second kind of reaction, deamination, removes the amino group of the amino acid in the form of ammonia. In the liver, the oxidative deamination of glutamate results in α-keto-glutarate (a TCA cycle intermediate) and ammonia, which is converted into urea and excreted. Deamination reactions in other organs form ammonia that is generally incorporated into glutamate to generate glutamine, which is the main transporter of amino groups in blood. Hence, all amino acids through transamination/deamination reactions can be converted into intermediates of TCA cycle, directly or via conversion to pyruvate or acetyl-CoA (Figure 5).