Natural Selection
In Charles Darwin's 1859
seminal book, On the Origin of Species, he tried to answer the question
of how species originate. He saw a paradox. On the one hand,
all living organisms attempt to perpetuate their kind by producing many more
offspring than are necessary to maintain their numbers. Yet, the actual
size of natural populations usually remains relatively constant over
time. How could this be? Darwin's answer was that many of the
offspring do not survive to reproduce. This phenomenon can be illustrated
by considering the common housefly (Musca domestica).
Females lay up to 500 eggs at a time.
The eggs hatch into larvae which go through several molting stages and then
transform into pupae. Thirty-six hours after emerging from pupae, females are receptive for mating. Adult flies live 15-30 days, during which time, females lay
eggs repeatedly though they mate only once. Over a 4-5 month period, the
descendents of a single mating pair of house flies potentially could number 1920. If that actually occurred,
we very quickly would be up to our armpits in fly bodies all over the planet and
the piles would grow at a rapidly increasing rate. Fortunately, most fly
eggs, larvae, and pupae are killed by other insects and microscopic
parasites. This keeps the total fly population more or less constant over
time. Darwin surmised that the environment operated in a selective way,
reducing the number of poorer-adapted variants of a species while increasing
the proportion of better-adapted ones. This process became known as
natural selection.
Darwin correctly understood that natural selection is usually the most powerful mechanism of evolution. However, he did not fully comprehend how it operates. This was due to the fact that he was largely ignorant of the mechanisms of genetics. That knowledge mostly came after his time. We now know that natural selection's effect on individuals depends on their phenotypes which in turn are determined mostly by their genotypes. The environment ultimately selects individuals with the best suited genotypes to survive to reproduce. Those individuals who have more surviving offspring pass on more of their genes to the next generation. As a consequence, the gene pool frequencies shift in the direction of their more adaptive alleles. However, the alleles that provide an advantage now may not in the future as new environmental stresses appear. Natural selection acts as a constantly changing template in its selection of winners and losers. This introduces chance into the equation. It is largely a matter of luck in having the right combination of genes at the right time to survive as the environment changes. Extinction occurs if those genes are not present.
For natural selection to cause evolution, it must select for or against one or more of the genotypes for a trait. In the simple case of a trait that is determined by a single gene with two alleles, there are five combinations of genotypes that nature can select:
|
1. |
either homozygote (AA or aa but not both) |
|
2. |
both homozygotes (AA and aa) |
|
3. |
either homozygote and the heterozygote (AA and Aa or aa and Aa) |
|
4. |
the heterozygote (Aa) |
|
5. |
all alleles (AA, Aa, and aa) |
Selection Against One of The Homozygotes
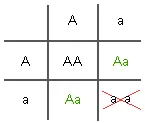
For traits that are controlled by a single gene that has two alleles, selection against one of the homozygotes (AA or aa) will result in a progressive decrease in the allele of which that unsuccessful homozygote consists. For example, if aa is completely selected against while AA and Aa are selected for, there will be only four possible successful mating patterns (as shown in the table below).
|
Selection against one of the homozygotes (aa) |
|||
|
Possible parent |
Expected offspring genotypes |
||
|
AA |
Aa |
aa |
|
|
AA X AA |
4 |
|
|
|
AA X Aa |
2 |
2 |
|
|
Aa X AA |
2 |
2 |
|
|
Aa X Aa |
1 |
2 |
1 |
|
Total |
9 |
6 |
1 |
Within one generation, the frequency of homozygous recessive (aa) children will drop dramatically. There will be a progressive decrease in the frequency of the "a" allele and a corresponding increase in the "A" allele every generation in which aa genotypes are selected against (as illustrated in the table below). This has been referred to as directional selection because of the shift in gene pool frequencies towards the advantageous allele.
|
Evolutionary trend
resulting from complete selection |
|||
|
Allele |
Generation |
||
|
1 |
2 |
3 |
|
|
A |
50% |
67% |
75% |
|
a |
50% |
33% |
25% |
However, the recessive allele (a) will not completely disappear since it is still passed on by heterozygous (Aa) parents to the half of their children who are likely to also be heterozygous.
|
When there is complete selection |
|
|
For the vast majority
of human genes, the pressure of natural selection is usually far more gentle. As a consequence, the
resulting evolution is so slow as to be difficult to detect in only a few
generations. In the case of recessive traits such as albinism  , homozygous recessive individuals are only at a slight selective
disadvantage. They usually live to adulthood and reproduce. In some
other genetically inherited recessive conditions, such as juvenile
onset diabetes , the selection has been more severe. In the
past, those who inherited it usually died in childhood before passing it on to
the next generation. As a result, the frequency of this recessive allele
was progressively reduced. This has all changed, however, since the
discovery of insulin in 1921. Diabetes is no longer the killer of
children it once was, and diabetic children grow up to have children with a
higher than average chance of inheriting this disease.
, homozygous recessive individuals are only at a slight selective
disadvantage. They usually live to adulthood and reproduce. In some
other genetically inherited recessive conditions, such as juvenile
onset diabetes , the selection has been more severe. In the
past, those who inherited it usually died in childhood before passing it on to
the next generation. As a result, the frequency of this recessive allele
was progressively reduced. This has all changed, however, since the
discovery of insulin in 1921. Diabetes is no longer the killer of
children it once was, and diabetic children grow up to have children with a
higher than average chance of inheriting this disease.
In the mid 1990's, a striking example of intense selection against one of the homozygotes for a trait came to light. This stemmed from the discovery that some people do not get AIDS even if they are repeatedly exposed to the HIV virus that is responsible for this usually fatal disease. The people who are immune have inherited two copies of a rare mutant gene known as CCR5-delta 32 --they are homozygous. Those who are heterozygous apparently have a partial immunity or at least a delay in the onset of AIDS. Approximately 10% of Europeans now have the CCR5-delta 32 gene variant, but it is extremely rare or absent in other populations of the world. There is a surprising connection in this story. The CCR5-delta 32 gene also provides immunity to a deadly disease of bacterial origin, bubonic plague . People who are homozygous for the CCR5-delta 32 gene variant are completely immune, while heterozygotes have partial immunity. It is very likely that this life-saving allele occurs as a random mutation and that it was selected for by the devastating black plague epidemics that swept over Europe beginning in the 14th century. During the first wave of plague, between 1347 and 1350, one fourth to one third of all Europeans died from this disease. Natural selection favored those who by chance had inherited the CCR5-delta 32 gene variant. Repeated waves of plague over the next three centuries resulted in an increase in the frequency of CCR5-delta 32 in the European population.
Because the CCR5-delta 32 gene variant has been found in the DNA of bones from some Europeans who lived more than 2,000 years before the medieval plague epidemics, it has been suggested that this gene may also have been selected for by other deadly contagious diseases such as smallpox.
Selection Against Both Homozygotes
If there is complete selection against both homozygotes (AA and aa) in childhood, the only possible mating will be between heterozygous individuals (Aa) because they will be the only ones who live long enough to reproduce.
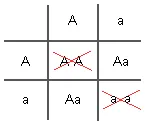
Extreme environmental conditions selecting only for heterozygous individuals can result in a balanced polymorphism in one generation. That is to say, the frequency of the two alleles (A and a) can each reach 50% and remain at that level so long as there is this sort of harsh natural selection. This has been referred to as stabilizing selection, or balancing selection, because there is not a shift in the gene pool frequencies towards one of the alleles.
|
Selection against both homozygotes (AA and aa) |
|||
|
Possible parent |
Expected offspring genotypes |
||
|
AA |
Aa |
aa |
|
|
Aa X Aa |
|
2 |
|
|
Total |
0 |
2 |
0 |
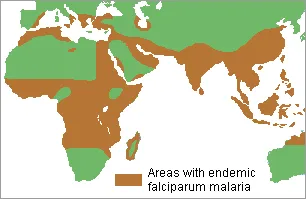
An example of nature
selecting against both homozygotes was found in Central Africa. This is an area
in which malaria has long been a serious problem. While
10% of the world's human population is infected by malaria, 90% of the
cases are in sub-Saharan Africa. It is the major cause of death
there. Children and pregnant women are especially vulnerable. An
African child dies of malaria every 30 seconds on average. Malaria is
caused by several related parasitic microorganisms (plasmodia ) that feed on red blood cells. The microorganisms are
transmitted from person to person by mosquitoes when they suck blood from their
victims. Mosquitoes do not get malaria
themselves. They only transmit the plasmodia along with small amounts of
the blood from their earlier victims. People who
produce normal red blood cells are good hosts and easily get the disease, which is
debilitating and ultimately often results in death.
|
Malaria caused by the |
|
|
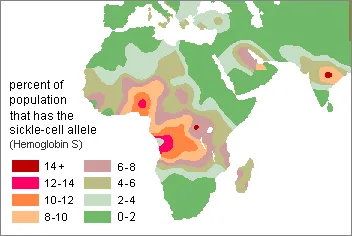
There is a high frequency of an inherited condition known as sickle-cell trait in African malarial zones. Homozygous recessive sicklers (aa) have resistance to falciparum malaria because their misshapen, deflated red cells are poor hosts. Unfortunately, these individuals usually die in childhood from bacterial infections made worse by weakened immune systems and severe anemia. About 100,000 people around the world succumb to sickle-cell related health problems every year. However, that is far fewer than the1,500,000 who die from malaria.
|
The sickling allele is at |
|
|
People who are heterozygous (Aa) for sickle-cell trait also have moderately good resistance to malaria because some of their red cells are misshapen and deflated, but they rarely develop the severe life threatening anemia and related problems typical of homozygous (aa) sicklers. Those who are homozygous dominant (AA) produce normal red blood cells, which makes them excellent hosts for malaria. Therefore, in falciparum malarial environments, nature selects for heterozygous sicklers. At the same time, it selects against homozygous sicklers and people who produce normal red blood cells.
|
|
|
|
|
Normal human red
cells |
Deflated red cells
from |
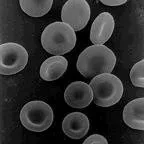
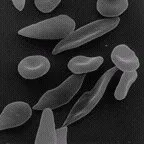
The sickling allele was not produced by natural selection. It apparently pops up periodically as a random mutation. Unless it is selected for, its frequency remains very low within a population's gene pool because it results in a selective disadvantage for those who inherit it. The presence of widespread falciparum malaria changes the situation. The otherwise harmful sickling allele provides an advantage for heterozygous individuals.
Selection favoring the sickling allele is an example of biocultural evolution . Human culture altered the environment, which resulted in factors that were advantageous to both the malarial microorganisms and the mosquitoes that transmit them between people. The sequence of events apparently began about 2000 years ago with the introduction into Africa of Southeast Asian root and tree crops that were adapted to the humid tropics. This resulted in an agricultural revolution and a subsequent human population explosion in sub-Saharan Africa. Slash-and-burn forest clearance for preparing agricultural fields altered the natural environment in a way that selected for the Gambiae group of anopheles mosquitoes that are largely responsible for spreading malaria in Africa. At the same time, the progressively increased density of humans made it easier for mosquitoes to find hosts and to inadvertently spread malaria. The more people who acquired malaria, the more likely it was for mosquitoes to transmit the malaria plasmodia to new hosts. Subsequently, the sickling allele became increasingly valuable as a population defense against the devastating effects of malaria. This natural selection by malaria in sub-Saharan Africa was not so complete as to result in a balanced polymorphism in just one generation. In fact, after nearly 2,000 years of selecting for the sickle-cell allele, it is not found to be above 40% in any major African population and is more often around 20%.
Sickle-cell trait is rare in North America with a single exception--African Americans. One in 12 of them carry the sickling allele and about 72,000 have sickle-cell anemia and other related clinical symptoms. One in 375 African American children is homozygous recessive for it. This is not surprising because most African Americans have ancestors who came from the malarial zones of West and Central Africa. Those who are heterozygous for this trait usually do not experience sickling problems unless they are exposed to a low oxygen environment for a prolonged period or do unusually strenuous physical activity. People with Hispanic, Mediterranean, or South Asian ancestry also have a somewhat elevated risk for inheriting the sickle-cell trait, but it is not as high as for Africans and their descendents. During the first decade of this century, 16 division 1 university football players in North America died suddenly without experiencing physical trauma beforehand. Nine of those men had a single copy of the sickling allele. It is likely that this contributed to their deaths.
Several other genetically inherited conditions may provide a degree of immunity to malaria in regions of the world in which sickle-cell trait is rare. Thalassemia and glucose-6-phosphate dehydrogenase deficiency (G6PD) may be in this category. They occur especially among people in South Asia and around the Mediterranean Basin. Both of these conditions result in severe anemia. As in the case of sickle-cell trait, this anemia apparently makes the victims poor hosts for the malaria plasmodia. It is likely that all three of these inherited blood abnormalities are biological solutions to the problem of surviving the harsh natural selection caused by malaria.
Selection Against The Heterozygote And One Of The Homozygotes
If natural selection is against an allele in both homozygous and heterozygous genotypes, the rate of change in gene pool frequencies will usually be much more rapid. In fact, it can result in the elimination of the targeted allele in only one generation. For example, if both aa and Aa genotype individuals fail to reproduce, then only AA people will contribute their genes to the next generation--the descendents will only inherit "A" alleles. This is an extreme form of directional selection.
|
Selection against the heterozygote and one of the homozygotes (Aa and aa) |
|||
|
Possible parent |
Expected offspring genotypes |
||
|
AA |
Aa |
aa |
|
|
AA X AA |
4 |
|
|
|
Total |
4 |
0 |
0 |
There are at least 5,000 genetically inherited human abnormalities and diseases. Apparently, many, if not most, of them are caused by recessive alleles. Usually, these alleles are carried without symptoms by heterozygous (Aa) individuals and are only selected out of the gene pool when homozygous recessive (aa) children are born. In order for humanity to be quickly rid of these diseases, there would have to be selection against both the heterozygous and the homozygous recessive individuals. However, this extreme form of natural selection is very rare.
Genetic testing and counseling is now often aimed at discouraging heterozygous carriers of harmful recessive alleles from reproducing. Sickle-cell trait and Tay-Sachs disease have been the main focus of this health campaign in North America. It has been particularly effective among the Ashkenazi (Eastern European) Jewish population in the United States. While they have a very high frequency of the allele for Tay-Sachs disease, the number of their children now born with it is low due to hard personal decisions that many Jews make based on genetic testing and education. By choosing not to have children, people who carry the recessive allele do not pass it on. The attempt at eliminating sickle-cell trait has been somewhat less successful among African Americans.
Selection Against The Heterozygote
When natural selection is only against heterozygotes, there will be four successful mating patterns (as shown in the table below). These will normally result in half of the children being heterozygous (Aa).
|
Selection against the heterozygotes (Aa) |
|||
|
Possible parent |
Expected offspring genotypes |
||
|
AA |
Aa |
aa |
|
|
AA X AA |
4 |
|
|
|
AA X aa |
|
4 |
|
|
aa X AA |
|
4 |
|
|
aa X aa |
|
|
4 |
|
Total |
4 |
8 |
4 |
However, if natural selection eliminates heterozygotes in childhood, the adult reproducing population will be genetically polarized. Half will normally be homozygous dominant (AA) and half will be homozygous recessive (aa) (as shown in the table below). This has been referred to as disruptive selection because both extremes are favored.
|
Selection against the heterozygotes (Aa) |
|||
|
Possible parent |
Expected offspring genotypes |
||
|
AA |
Aa |
aa |
|
|
AA X AA |
4 |
|
|
|
AA X aa |
|
|
|
|
aa X AA |
|
|
|
|
aa X aa |
|
|
4 |
|
Total |
4 |
0 |
4 |
Selection Against All Genotypes
When nature completely selects against all genotypes (AA, Aa, aa), the result is that neither of the two alleles will appear in the next generation. More importantly, extinction of the population will occur since all genotypes are at a selective disadvantage.
Complications of Natural Selection
In all of the natural selection examples given so far, it has been assumed that there are only two alleles of each gene. However, some traits are controlled by many more alleles. In addition, simple Mendelian rules of dominance do not always hold, especially in the case of polygenic traits. It must be assumed that the way in which nature selects for or against such traits can be more complex than described here.
Through culture created technology, modern humans have been able to alter selective pressure for or against certain genes. This is mainly a consequence of two actions that are having a profound impact on our human gene pool. We have very likely increased the rate of mutation by inadvertently releasing many mutagenic chemicals and radiation into our environment. At the same time, modern medicine has reduced discrimination against harmful disease causing genes by developing cures for what previously had been fatal conditions. In the past, these genes were generally weeded out of our gene pool by natural selection killing off those who carried them. A form of eye cancer in young children, known as retinoblastoma, provides an example of this phenomenon. It is a rare disease, affecting only about 4 out of every million babies. Since it is a dominant trait, both homozygous dominant and heterozygous genotypes result in retinoblastoma. Prior to the development of surgical procedures to treat this cancer, it was virtually always fatal. Its victims died before they could pass it on to another generation. Most new cases were probably the result of extremely rare mutations in sex cells. With adequate treatment, 70% of the patients can now survive retinoblastoma and can transmit it to at least 50% of their offspring. As a result of preventing children from suffering and dying from a horrible disease, we have increased the likelihood that more children will be born with it. This will make us ever more dependent on new costly medical treatment procedures. Is this wrong? Most of us would say of course not. Morally we cannot do otherwise. We cannot let children die if we can prevent it. However, it does create a growing dilemma for humanity. We potentially are building up an ever larger genetic load of harmful genes because we have hindered nature's ability to eliminate them from our gene pool.